
Open Journal of Medical Psychology > Vol.7 No.4, October 2018
Psychobiological Model of Bipolar Disorder: Based on Imbalances of Glial-Neuronal Information Processing
Bernhard J. Mitterauer
Volitronics Institute for Basic Research, University of Salzburg, Salzburg, Austria.
DOI: 10.4236/ojmp.2018.74008 PDFHTML XML 1,057 Downloads 2,345 Views Citations
A psychobiological model of the etiopathology of bipolar disorder is proposed. Based on genetic-epigenetic and chronobiological factors a hyperintentional personality structure, if faced with non-feasible intentional programs in the environment, suffers from inner and outer stress. This stress situation leads to imbalances in information processing in glial-neuronal synaptic units, called tripartite synapses. In depression the overexpression of astrocytic receptors and of gap junctions in the astroglial network causes a prolonged information processing which affects the behavior generating systems in the brainstem reticular formation. Because the activation of the behavior generating systems is protracted, they are unable to select an appropriate mode of behavior (e.g. communicating, eating, working, sleeping, etc.) from sensory information in real time. Inversely, in mania astrocytic receptors and gap junctions are underexpressed causing a shortened synaptic information processing with rapid changes in behavior. Switching may represent a coping-attempt with depression by mania and vice versa. Towards a comprehensive model of the pathophysiology of bipolar disorder the role of microglia and their devastating effects on glial-neuronal interactions are outlined. Finally, the testing of the model is discussed.
Keywords
Bipolar Disorder, Glial-Neuronal Interactions, Synaptic Imbalances, Hyperintentionality
Share and Cite:
Mitterauer, B. (2018) Psychobiological Model of Bipolar Disorder: Based on Imbalances of Glial-Neuronal Information Processing. Open Journal of Medical Psychology, 7, 91-110. doi: 10.4236/ojmp.2018.74008.
Bipolar disorder is a life-long affective disorder affecting approximately 2% – 5% of the world’s population [1] . Generally, mood disorders are best characterized as syndromes rather than disease entities [2] . Bipolar disorder represents a special syndrome consisting of states of depression or mania with polar-opposite syndromes. Periods between these extreme states represent euthymic states. In addition, mixed manic-depressive states can also occur.
Typical symptoms of mania are elevated mood, grandiosity, risky behaviors, reduced sleep and racing thoughts. In contrast, typical symptoms of depression are depressed mood, anhedonia, worthlessness, difficulty in thinking and concentration, insomnia or hypersomnia [3] . In states with mixed features the prevailing mood is dysphoria [4] . The etiopathology of bipolar disorder is still unknown. Explanatory models focus on biological, psychological and sociological approaches. Current biological hypotheses comprise changes in genetics, systems of neurotransmitter and neurotrophic factors, neuroinflammation, stress system activity, chronobiology and mitochondrial dysfunctions [5] . In addition, psychosocial factors mainly concern trauma, negative life events, social support deficits and family problems [6] . Basically, bipolar disorder is multifactorially determined caused by stress-diathesis interactions as described in the final common pathway model of Akiskal and McKinney [7] . Towards a “unified field theory” Maletic and Raison [8] elaborated on an etiopathophysiological model of bipolar disorder. Basically, the etiology described by these researchers comprises genetic-epigenetic factors and stress causing dysregulations and structural changes in the neuro-glial networks. In addition, circadian dysregulations and dysregulations of the immune system at the cellular and subcellular level are responsible for the clinical symptomatology of bipolar disorder.
Although most hypothetical models of depression and bipolar disorder are neurocentric, experimental findings indicate that the glial system, especially astrocytes and microglia, plays a significant role in the pathophysiology of affective disorders [9] [10] [11] . Basically, the brain consists of the neuronal cell system and the glial cell system. Ca2+ imaging and electrophysiological techniques support the existence of a complete bidirectional communication between astroglia and neurons, and indicate an important active role [12] exerting a modulatory function in synaptic information processing [13] . Therefore, the pathophysiological model of bipolar disorder here proposed is based on glial-neuronal interactions focusing on imbalances in astrocyte-neuronal synaptic units, called tripartite synapses [14] [15] [16] [17] .
Normally, a person is intending to realize his (her) programs (goals, wishes, desires). If intentional programs are not feasible in the environment and the person persists on realizing them, then the psychobiological state is hyperintentional striving for the realization of the unfeasible. Here intentionality is defined as follows: an intentional program generates a specific multirelational structure in the inner and outer environment based on the principle of that program [18] . My clinical observation and treatment of patients suffering from bipolar disorder over the years led me to the conception of a hyperintentional personality structure of patients genetically-epigenetically inclined to bipolar disorder.
The model is this: the character feature of hyperintentionality may cause imbalances of information processing in glia-neuronal synaptic units, called tripartite synapses. A tripartite synapse consists of the presynapse and the postsynapse as the neuronal component and the astrocyte and its network as the glial component [19] . I hypothesize that astrocytic receptors ready for occupancy by activating substances (neurotransmitters, neuromodulators etc.) cannot exert their balancing function. Based on a formalism of balancing it can be shown, if a system is balanced, underbalanced or overbalanced. If in a depressive state the synaptic information processing is underbalanced, since astrocytic receptors outnumber the amount of activating substances, the system may develop depression. In a manic state astrocytic receptors are underexpressed so that a surplus of activating substances arises generating an overbalanced synaptic information processing [16] [17] . In the case that the receptors in tripartite synapses are both underexpressed and overexpressed in different brain regions a mixed manic-depressive state becomes generated. The overexpression of astrocytic receptors leads to a relative lack of neurotransmitter substances that cause a prolonged information processing in depression. Inversely, if astrocytic receptors are underexpressed, transmitter substances overactivate astrocytic receptors leading to a shortened synaptic information processing in mania.
Underbalance and overbalance of synaptic information processing also affect the generation of the normal modes of behavior such as eating, working, sleeping, communicating etc. dependent on prolonged or shortened information processing. If information processing is prolonged, the behavior-generating systems in the brain stem reticular formation or in the brain stem core systems cannot process environmental information in real time causing the persistence of one or more modes of behavior observed as retardation in depression. In the case of shortened information processing environmental information becomes rapidly processed, responsible for a highly irritable and inconstant behavior in mania.
Fundamentally, the pathophysiological dysfunctions and structural changes described may generate a hyperintentional system unfeasible in depression. The switch to a manic state can be interpreted as a “coping-attempt” with the unfeasible, since information becomes rapidly processed so that thoughts and actions make one feel sure that “all is possible” what I called pseudo-omnipotence [20] .
3. Hyperintentional Personality Structure
Long-term therapeutic communication with patients suffering from bipolar disorder led to the concept of hyperintentional personality structure as a main psychological factor of inclination to depression or bipolar disorder [14] [15] . Generally, an intentional program generates a specific multirelational structure in the inner and outer environment based on the principle of that program [18] . Hyperintentionality occurs, if an intentional program persists, despite it being non-feasible in the environment. One can also speak of high aspirations of patients inclined to affective disorders [21] .
Parents of children susceptible to bipolar disorder are convinced of having a genius daughter or son who will do great things and innovations in the future. Genius artists and scientists after a high creative period are suddenly incapable of working without a conceivable reason. We treated a great computer scientist who invented a novel computer system in a sleepless working rush, but he became more and more unable to communicate his system with engineers for technical implementation since he doubted the novelty of the invention.
Hyperintentionality of persons susceptible to bipolar disorder may be basically determined by an epigenetic process and education, in which certain genes are expressed in parent-origin-specific manner. Basically, epigenetics attempts to provide a framework for understanding how the expression of genes is influenced by experience and environment [22] , personality [23] , and mental disorders [24] or affective disorders [25] .
Interestingly, Freud [26] described mental disorders as narcissistic neuroses. Currently, the concept of narcissism has been introduced in psychology and psychopathology characterizing absolute egocentric personalities, but the original meaning of narcissism in the poem of Ovid [27] is not only absolute self-reference or egocentricity, but represents basically non-feasible intentionality. Narcissus strives to meet and touch his double, but he fails to grasp his mirror image in a pond and deceases. Significantly, the repeated and rapid grasping for the mirror image is generating water waves which make the mirror image “fluid”. Hence, the attempts of Narcissus to “comprehend” himself is an interplay between grandiosity and self-destruction characteristic of a manic-depressive behavior.
4. The Tripartite Synapse and the Astroglial Network
The close morphological relations between astrocytes and synapses as well as the functional expression of receptors in the astrocytes led to the new concept of the tripartite synapse [19] . Arague and colleagues showed that astrocytes respond to neuronal activity (neurotransmitters) with an elevation of their internal Ca2+ concentration which triggers the release of chemical transmitters (gliotransmitters) from astrocytes themselves, and, in turn, causes feedback regulation of neuronal activity and synaptic strength.
Astrocytes are interconnected by gap junctions (GJ) forming an astroglial network. GJs are composed of hemichannels (connexons) that dock to each other via their extracytoplasmic extremities. Basically, GJs are considered to provide a structural link by which single cells are coupled to build a functional network with a communication behavior that cannot be exerted by individual cells. GJs of an astroglial network consist of different kind of connexin proteins (Cx) forming homotypic (same connexins) or heterotypic (different connexins) gap junction channels. The connexin biosynthesis is dynamically regulated so that GJs can also be under or overexpressed [28] . My model of affective disorders is based on the hypothesis that the overexpression and underexpression of connexins does not only dysregulate the astroglial network, but may also influence the expression of astrocytic receptors.
Figure 1 shows a model of a glutamatergic tripartite synapse and the astroglial network. The neurotransmitter glutamate (GLU) is released from the presynapse activated by a dendrite (D). GLU occupies postsynaptic receptors (por) and receptors on the astrocyte (acr). Transporters (t) reuptake GLU on the presynapse again. The activation of acr leads to an increase of Ca2+ concentration and the production of gliotransmitters (GT). GTs released from the astrocyte activate presynaptic receptors (psr), por and extrasynaptic receptors (esr). The occupancy of psr by GT exerts a negative feedback mechanism temporarily turning off neurotransmission. The astrocyte is interconnected with neighboring astrocytes via gap junctions (GJ) building an astroglial network.
5. Balance and Imbalances of Synaptic Information Processing
The interaction between neurotransmitters (NT) and the astrocytic receptors (acr) can be system-theoretically interpreted [29] as balanced, underbalanced,

Figure 1. Model of a glutamatergic tripartite synapse and the astroglial network. Activated by a dendrite (D) glutamate (GLU) is released from the presynapse in the synaptic cleft (SC). GLU activates postsynaptic receptors (por) and is reuptaken on the presynapse by transporters (t). In parallel, GLU occupies receptors on the astrocyte (acr) activating channels which lead to an increase in calcium concentration (Ca2+↑) and the production of gliotransmitters (GT). The release of GT from the astrocyte activates presynaptic receptors (psr), por and extrasynaptic receptors (esr) on the postsynapse. The effect of GT corresponds with a negative feedback mechanism on the presynapse and the depolarization by the occupancy of por and esr. The astrocyte is interconnected with the astroglial network (double headed arrow) that is built by gap junctions (GJ, small squares).
overbalanced, or even unbalanced. Formally, neurotransmitters represent values and astrocytic receptors variables. In a balanced system the number of variables (acr) and the number of values (NT) is equal. If the variables (acr) dominate the values (NT) available in the system, the system is underbalanced, responsible for depression. Inversely, if the values (NT) dominate the variables (acr) the system is overbalanced, which may occur in manic states [16] . In the case that no appropriate variables (non-functional acr) are available, the system is totally unbalanced. This system state may be responsible for the pathophysiology of schizophrenia [30] .
In Figure 2 is the formal principle of system-balancing signified by two inverse parallel lines. The length of the line above indicates the concentration of

Figure 2. Balance, imbalance, and unbalance between neurotransmitters (NT) and astrocytic receptors (acr). The concentration of NT and the amount of acr is signified by two inversive parallel arrowed lines. If the concentration of NT (values) for the occupancy of acr (variables) is appropriate, the synaptic system is in balance (a). In the case of an excess of acr, the amount of NT is too small, leading to an underbalanced synaptic system (b). This state may cause depression. If the concentration of acr is low in relation to a normal NT concentration, the synaptic system is overbalanced (c). Overbalanced synaptic states may cause mania. Supposing that no appropriate acrs are available (dashed line) the system is totally unbalanced (d). This synaptic state may be responsible for the pathophysiology of schizophrenia.
neurotransmitters, the length of the line below stands for the amount of astrocytic receptors. Imbalances are signified by shortened or dashed lines.
6. Underbalanced Tripartite Synapses May Cause Depression
Figure 3 outlines an underbalanced tripartite synapse, since gap junctions in the astroglial network and astrocytic receptors are overexpressed. The upregulation of connexins may cause an overexpression of astrocytic receptors which cannot be occupied by neurotransmitters (NT) in real time. This causes an underproduction of gliotransmitters (GT) because Ca2+ concentration is diminished so that GTs negatively feedback to the cognate presynaptic receptors and synaptic information processing is protracted. In addition, the reuptake of NT decreases the amount of NT in the synaptic cleft. Such delay of information processing could explain the main symptoms of depression such as feelings of insufficiency, disturbances of circadian rhythms and hyperintentionality [31] .
Importantly, Quesseveur and colleagues [32] investigated the role of hippocampal astroglial connexin 43 in emotionality and the effects of selective serotonin reuptake inhibitors. Given that phosphorylation is a prerequisite for acute function of connexins [33] , the therapeutic effects of antidepressant drugs might implicate the functional inactivation of connexin 43. This effect may support my model of the pathophysiology of depression, since it allows the interpretation that overexpressed connexins become reduced by antidepressant drugs balancing

Figure 3. Upregulation of connexins and astrocytic receptors responsible for depression. Neurotransmitters (NT) released from the presynapse activate postsynaptic receptors (por) and astrocytic receptors (acr). The upregulation of glial connexins forming gap junctions (GJs, red) upregulates the expression of acr. Overexpressed acr cannot completely be occupied by NT causing a diminished concentration of Ca2t and a prolonged production of gliotransmitters (GT) (dashed lines). This leads to a protracted activation of psr, por and extrasynaptic receptors (esr) so that neurotransmission is delayed [17] [31] (SC: synaptic cleft).
synaptic transmission. Moreover, functional imaging studies of patients with depression indicate that the clinical response to antidepressants is different in the hippocampal and prefrontal region. Although antidepressant substances reduce functional activity in the hippocampus and the limbic regions, they increase functional activity in the prefrontal cortex [34] . I suggest that in depression the overexpression of gap junctions and astrocytic receptors operate hyperintentionally in emotion processing regions such as in hippocampus. Since in cognition processing areas as the prefrontal cortex, gap junctions and astrocytic receptors are underexpressed, environmental information cannot be processed in time “flooding” synaptic information processing in the sense of cognitive irritability.
Most of the typical receptors for neurotransmission have been identified on the astrocytic membrane [35] . Although dysregulations of the expression of astrocytic receptors are as yet not elucidated in depression, upregulations of astrocytic receptors are already found in Alzheimer’s disease [36] and in Parkinson’s disease [37] . Importantly, the Fuxe Group investigates receptor-receptor coupling in tripartite synapses and showed that abnormal receptor-receptor coupling in inflammation, possibly responsible for depression, dysregulates neurotransmission [38] . Basically, our understanding of the functions and dysfunctions of receptors in synapse-astrocyte interactions could elucidate the role of astrocytic receptors in depression and mania.
One may argue that the decrease and hypofunction of astrocytes seems to speak against the model proposed here. However, there is some evidence that receptors on astrocytes are upregulated. Animal models of chronic stress show that adenosine A2A receptors are upregulated and polymorphisms of A2A receptors are associated with emotional disturbances and their over-expression triggers emotional dysfunction [11] . This mechanism could also operate in depression and should be elucidated in brains with depression.
7. Overbalanced Tripartite Synapses May Cause Mania
Whereas synaptic information processing in depression is protracted, a manic state may be generated by inverse mechanisms which shorten information processing. Basically, the reduced number of gap junctions in the astroglial network (syncytium) and the underexpression of astrocytic receptors cannot exert a balancing function in tripartite synapses. Under these conditions there will be a surplus of neurotransmitters relative to the underexpressed astrocytic receptors. In parallel, astrocytic receptors are flooded with transmitters. This flooding also influences the negative feedback mechanism on the presynapse with the effect of shortened cycles of information processing (Figure 4).
Because the glial intentional programs embodied in the astrocytic receptors are immediately realizable and “all seems to be appropriate”, a manic patient feels grandiosity which I call pseudo-omnipotence [20] . Dependent on the transmitter systems or brain areas affected, the synaptic overbalance may be
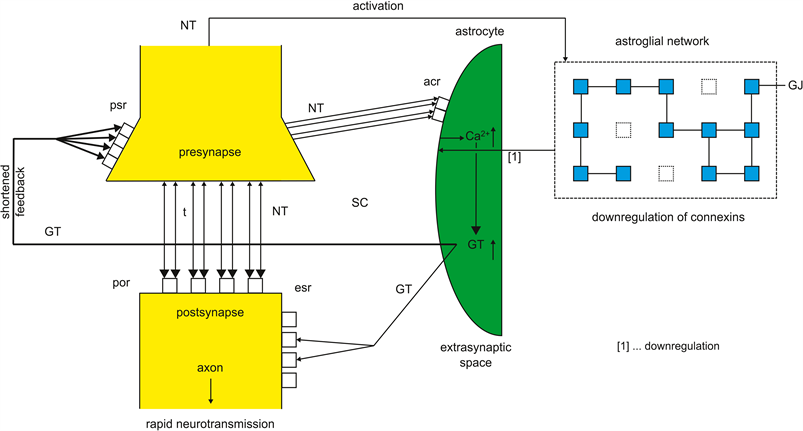
Figure 4. Overbalanced tripartite synapse responsible for mania. Neurotransmitters (NT) released from presynapse activate postsynaptic receptors (por) and astrocytic receptors (acr). The downregulation of glial connexins forming gap junctions (GJ) downregulates the expression of acr. Underexpressed acr are flooded by NT causing an increased Ca2+↑ concentration and an increasing production of gliotransmitters (GT↑) in the astrocyte leading to a shortened feedback on presynaptic receptors (psr) (bold line). The reuptake of NT on the presynapse by transporters (t) does not significantly decrease the amount of NT in the synaptic cleft (SC). The activation of extrasynaptic receptors (esr) is also intensified causing a rapid neurotransmission.
responsible for the pathophysiology of manic symptoms such as euphoria and feelings of grandiosity. In addition, the rapid synaptic cycles could be explanatory of manic distractibility, flight of ideas, overactivity and circadian and biorhythmic disturbances, especially insomnia.
8. Overbalanced and Underbalanced Tripartite Synapses May Cause Mixed Manic-Depressive Episodes
Momentary tearfulness, depressed mood, suicidal ideation are commonly observed at the height of mania or during the transition from mania to depression. Another common feature is racing thoughts in the context of retarded depression. Those transient labile periods must be contrasted with mixed episodes in the long-term course of bipolar disorder. Mixed episodes are characterized by dysphorically excited moods, anger, panic attacks, persevered speech, agitation, suicide ideation, severe insomnia, grandiosity, hypersexuality, persecutory delusions and confusion [2] . Importantly, diagnostics of mixed states is broadened in the Diagnostic Statistical Manual (DSM-5) [4] .
If a patient shows depressive and manic symptoms at the same time, it seems likely that synaptic information processing is in some brain areas overbalanced while underbalanced in other areas as outlined in the Figure 3 and Figure 4. Approximately 40% of bipolar patients suffer from mixed states. Current hypotheses or models refer mainly to circadian genes, neurotransmitters and fluctuations in affective states. In the perspective of imbalances of astrocyte-synapse interactions underbalanced and overbalanced synaptic systems may operate in mixed episodes. Although this pathophysiological model is as yet not experimentally tested, relevant studies indicate that mixed states result from genetic susceptibility in the circadian and dopamine neurotransmission and dysregulations in the catecholamine-acetylcholine neurotransmission balance which leads to mood fluctuations [8] [39] .
9. Switching from Depression to Mania May Represent a Coping-Attempt with Depression
Although the switching mechanism is still not fully explored, consistent experimental findings provide evidence that dysregulations of glial-neuronal interactions with abnormalities in glial elements are more apparent than in neurons [11] . This means that not only astrocytes and their networks are affected, but also microglia and oligodendroglia with myelin. In the model here proposed permanent stress is generating dysfunctions and structural impairments, especially in the glial system. In addition to environmental stress in a broad sense, inner stress is caused by imbalances of information processing activating microglia with structural changes in the whole glial system [40] .
Importantly, the hypothesis of dopaminergic dysfunction in bipolar disorder may support the pathophysiological model outlined. Accordingly, switching occurs, if excessive dopaminergic activity in the course of mania down-regulates dopamine receptors which trigger a transition into a depressive state [41] . In an overbalanced tripartite synapse astrocytic receptors are underexpressed leading to shortened information processing and flooding of receptors with neurotransmitters. Although various receptor and transmitter systems may be affected, excessive dopaminengic activity represents a typical experimental finding. However, the excess of dopaminengic transmission may be basically caused by the underexpression of astrocytic receptors as depicted in Figure 4.
Importantly, neurotransmitter flooding generates an increasing inner stress which forces the synaptic system to switch to inverse pathophysiological mechanisms slowing down synaptic information processing responsible for depression. On the contrary the switch from depression to mania could represent a coping-attempt with depression. The rapid information processing in a manic state makes the patient convinced of being highly potent in cognition and action, but unable to reflect his (her) mostly destructive behavior regarding to him (her) self and to the society.
10. Displacement of Action Pattern Generation in the Brain Stem Reticular Formation
On the behavioral level animals or humans must not only select action programs adapting to the environment situation, but they are also able to intend an action in the environment. Basically, one of the most fundamental principles in system dynamics is the premise that the structure of the system will generate its behavior, commonly interpreted as modes of behavior [42] . The reticular formation in the brain stem is a candidate system for generating the modes of behavior such as eating, working, sleeping, communicating etc, since the reticular formation is interconnected with all regions of the brain [43] [44] including life sustaining nuclei that command and control e.g. heart beat and inspiration [45] [46] .
Figure 5 shows a model of the generation of six modes of behavior in time periods t1∙∙∙t6. The time periods t1∙∙∙t6 correspond to the action programs a1∙∙∙a6 and represents the period in which one of the six modes of behavior is activated. Environmental information activating the perception systems is processed in synapses of special cortical and subcortical systems [47] . This figure refers to the cognitive, emotional, psychomotor and autonomic-circadian systems. Dependent

Figure 5. Model of the generation of modes of behavior in the period t1∙∙∙t6. Environmental information activates the perception systems (one headed arrow). This information is processed in synapses of special cortical and subcortical areas. Cognitive, emotional, psychomotor and autonomic-circadian systems activate pertinent action programs (a1∙∙∙a6) in the brainstem reticular formation. At the moments t1∙∙∙t6 the reticular formation selects most appropriate information to an action program a1∙∙∙a6 such as mental activity, anxious, euphoric, work, eat, sleep (double headed arrows signify bidirectional interactions).
on the action program activated by synapses, the integrative function of the reticular formation is capable of commanding and controlling the generation of a pertinent mode of behavior such as mental activity, feeling of anxiety, euphoria, working, eating, and sleeping [48] .
In a depressive state the protracted information processing in tripartite synapses may cause a displacement of the generation of the modes of behavior which we could demonstrate in a computer simulation of hypersomnia [15] . In the case of a manic state information processing is shortened leading to rapid changes of action patterns which we observe as an irritable behavior of these patients.
Principally, the life-long affection with depressive and manic episodes in various courses may be caused by the persistence of the hyperintentional personality structure. Whereas in euthymic states stress situations may not activate genes responsible for the pathophysiology of depressive or manic episodes, the persistence of intentional programs striving for their feasibility can further elicit imbalances and structural changes of information processing as outlined in the present model of bipolar disorder.
11. Towards a Comprehensive Model of the Etiopathology of Bipolar Disorder
Although the present study focuses on the central role of astrocytes and their networks in synaptic imbalances, a comprehensive model of pathophysiology of bipolar disorder must also refer to microglia and oligodendroglia. Basically, all glial cells are interconnected via gap junctions building an active system in its interactions with the neuronal system [49] . Any pathological event in the brain leads to the activation of microglia, the immunocompetent cells of the central nervous system [50] . Experimental findings suggest that active microglia and increased proinflammatory cytokines play an important role in the pathophysiology of bipolar disorder with a possible link between neuroinflammation and peripheral toxicity [51] [52] .
As already described hyperintentionality exerts chronic stress that not only activates microglia, but also triggers microglia dystrophy [40] . Inflammatory substances released by microglia initiate a positive feedback loop in which astrocytes also begin to release adenosine-tri-phosphate (ATP) and cytokines, which trigger further inflammatory cytokine release from microglia, thus perpetuating the inflammatory cycle. Activated microglia reduces the release of neurotropic factors further perpetuating microglia activation leading to the impairment of oligodendroglia and neuronal apoptosis [53] . Importantly, the convergence of genetic vulnerabilities in bipolar disorder appears particularly target oligodendrocyte function [54] , but imaging studies of brains with bipolar disorder demonstrate structural impairments of all glial cell types [55] .
Considering the structural impairment of oligodendroglia it is of significance that their myelin sheaths are also destructed. Since myelin sheaths enable a saltatory axonic conduction making conduction faster, oligodendrocyte-myelin impairment slows down axonic conduction. Therefore, prolonged information transfer from synapses to the behavior-generating systems in the brain stem, responsible for depression, may be intensified by impairment or loss of oligodendroglia and myelin.
Family and twin studies suggested a strong genetic component in the etiology of mood disorders [56] . Although the inheritance of bipolar disorder seems to be beyond any doubt, the mode of transmission and the nature of the transmitted abnormalities are still not fully understood. We investigated a sample of 307 (N = 6704) sibling groups affected with bipolar disorder able to trace the pedigrees in some cases as far as six generations and found a significant genetic loading [57] .
Circadian dysregulation in bipolar disorder is experimentally well established. Mutations of clock genes may play a significant role [58] [59] [60] . Morningness and evening preference has been linked with bipolar disorder. The secretion of several neurotransmitters is regulated by circadian rhythm and is altered in bipolar disorder. In the perspective of synaptic imbalances described in the present model circadian dysregulation of neurotransmitter secretion may influence imbalances of synaptic information processing. For example, serotonin synthesis is controlled by significant diurnal and seasonal rhythmic fluctuations. In a preclinical model has been shown that serotonin influences the transcription of the CLOCK genes. However, the relationship between bipolar disease and the role of monamines in the disturbance of circadian regulation is as yet not fully understood [60] .
In Figure 6 a pathophysiological model of bipolar disorder is outlined. It is based on glial-neuronal interactions. Genetic-epigenetic factors [61] , circadian dysregulations [62] and stress determine dysregulations of glial-neuronal interactions. Stress activates microglia, which exerts functional dysregulations and structural changes of astroglia and oligodendroglia. In depression astrocytic receptors and glial gap junctions are overexpressed leading to a prolonged information processing (Figure 3). In mania astrocytic receptors and glial gap junctions are underexpressed causing a shortened synaptic information processing (Figure 4). Switching from depression to mania and vice versa represents an inverse operation as a coping-attempt with extreme pathophysiological imbalances and behavioral impairments. In a depressive state prolonged synaptic information processing leads to protracted axonic information transfer to the behavioral-pattern generation system in the brain stem reticular formation so that the modes of behavior cannot be selected in time and one or more modes persist in the sense of behavioral retardation. Inversely, in a manic state the very shortened synaptic information processing accelerate axonic information transfer causing a rapid change of the modes of behavior observed as manic irritability. All the system dysregulations generate chronic system stress that co-determines structural impairments or loss of cells.

Figure 6. Outline of a pathophysiological model of bipolar disorder based on imbalanced glial-neuronal interactions. Based on genetic-epigenetic factors and circadian dysregulations stress activates microglia (M) which causes functional dysregulation and structural changes of astrocytes and oligodendrocytes (oc) with myelin (My). In depression astrocytic receptors (acr) and glial gap junctions (GJ) are overexpressed leading to a prolonged synaptic information processing (dashed lines) (see also Figure 3). In mania acr and GJ are underexpressed causing a shortened synaptic information processing. Depressive states can switch (double headed arrow) to manic states and vice versa. In depression the prolonged synaptic information processing and the protracted axonic information transfer causes a persistence of one or more modes of behavior (fat rectangle). Inversely, in mania the shortened synaptic information processing leads to a rapid change of the modes of behavior (double headed arrows between squares). Cell decay is depicted (black circle).
Considering dopaminergic dysfunction in bipolar disorder it has been hypothesized that excessive dopaminergic activity in the course of mania precipitates dopamine receptor down-regulation, which subsequently triggers a transition into a depressive state [8] [41] . Importantly, this model only refers to the neuronal synapses and not explicitly to the receptors on the astrocyte. Moreover, if we assume that astrocytic receptors are down-regulated in a manic state leading to excessive dopaminergic activity in synaptic information processing, which subsequently triggers the upregulation of astrocytic receptors, then a relative lack of dopamine in the synaptic cleft causes a prolonged information processing. This imbalance may represent the basic disorder in depression. Inversely, if excessive dopaminergic activity caused by the underexpression of astrocytic receptors floods the synaptic cleft, a manic and not a depressive state becomes generated.
Currently, experimental findings indicate that receptor-receptor coupling plays a significant role in synaptic information processing [38] . These experiments may contribute to elucidate the role of astrocytic receptors in synaptic imbalances. Basically, my model of bipolar disorder is testable, but faced with limitations. Whereas the number of astrocytic receptors can be counted with in vivo imaging techniques [63] , the amount of over- or underexpressed gap junctions in comparison to the expression of gap junctions in the balanced network may be difficult to access because of the high complexity of astroglial networks [64] . In addition, hyperintentionality and its effects on the pathophysiology of bipolar disorder must be investigated. Here, animal models are inappropriate, since nobody knows, if the animal is in a hyperintentional state in the sense of a long-term intentional program. The capability to strive for the realization of long-term goals may be exclusively a human feature.
Although a hyperintentional personality structure can be assessed with a semi-structured interview, my pertinent experience with patients suffering from bipolar disorder demonstrates that the patients are often not aware of their persistent striving for an unfeasible goal. However, therapeutic communication and deep interviews over a longer time span mostly reveal the hidden burden of hyperintentionality.
The basic therapeutic implications from the model here proposed consist of both a long-term psychopharmacological relapse prevention and adjunctive psycho-social strategies [65] .
The significance of the present model lies in presenting a new psychobiological model of bipolar disorder based on synaptic imbalances of glia-neuronal interactions which are determined by a hyperintentional personality structure. The model focuses on the expression of astrocytic receptors and their role in balancing synaptic information processing. Based on a formalism of balance it is hypothesized that, if the number of astrocytic receptors corresponds to the concentration of neurotransmitters released from the presynapse, the neurotransmission is balanced. In the case that astrocytic receptors and the gap junctions in the astroglial network are overexpressed, neurotransmitters cannot activate all receptors in time. This system state is underbalanced leading to a prolonged neurotransmission responsible for depression. Inversely, if astrocytic receptors and glial gap junctions are underexpressed, a surplus of neurotransmitters arises causing a shortened neurotransmission. This synaptic state is overbalanced and responsible for mania. In mixed manic-depressive states tripartite synapses in some brain areas may be overbalanced and in other areas underbalanced.
In addition, my model suggests that the pattern of astrocytic receptors embodies an intentional program which modifies synaptic information processing. Since in depression overexpressed astrocytic receptors operate hyperintentional, the switching from depression to mania represents a coping-attempt with depression. Importantly, the main symptoms of depression and mania can be deduced from the synaptic imbalances described. In depression synaptic information processing is prolonged so that the behavior generating systems in the brain stem cannot generate the normal pattern of the modes of behavior (e.g. communicating, eating, working, sleeping, etc.) in real time. Consequently, one or more of the modes of behavior persist, typical for retardation or agitation on the behavioral level. Since in mania information processing is shortened, a rapid change of behavior occurs observed as manic irritability and flight of ideas.
Currently, experimental findings indicate that all main glial cells play a central role in the pathophysiology of mental or affective disorders [10] . Importantly, Maletic and Raison [8] elaborated an “integrated neurobiology of bipolar disorder” focusing on dysregulations of glial-neuronal interactions. Because of the heterogeneity of the course of illness and the high pathophysiological dynamics, these researchers challenge the diagnostics of the bipolar disorder and propose a new “unified field theory”. Working in this direction my model may represent a new approach to the research of affective disorders, since the psychological component of hyperintentionality is integrated in the pathophysiological mechanisms.
I am grateful to Christian Streili for designing the figures and tables, and Marie Motil for preparing the final version of the paper.
Conflicts of Interest
The authors declare no conflicts of interest.
| [1] | Clemente, A.S. (2015) Bipolar Disorder Prevalence: A Systematic Review and Meta-Analysis of the Literature. Revista Brasileira de Psiquiatria, 37, 155-161. https://doi.org/10.1590/1516-4446-2012-1693 |
| [2] | Akiskal, H.S. (1995) Mood Disorders: Introduction and Overview. In: Kaplan, H.J. and Sadock, B.J., Eds., Comprehensive Textbook of Psychiatry, Williams and Wilkins, Baltimore, 1067-1079. |
| [3] | Young, J.W. and Dulcis, D. (2015) Investigating the Mechanism(s) Underlying Switching between States in Bipolar Disorder. European Journal of Pharmacology, 759, 151-162. https://doi.org/10.1016/j.ejphar.2015.03.019 |
| [4] | American Psychiatric Association (2013) Diagnostic and Statistical Manual of Mental Disorders. 5th Edition, American Psychiatric Association, Washington DC. |
| [5] | Sigitova, E., Fisar, Z., Hroudová, J., Cikanková, T. and Raboch, J. (2017) Biological Hypotheses and Biomarkers of Bipolar Disorder. Psychiatry and Clinical Neurosciences, 71, 77-103. https://doi.org/10.1111/pcn.12476 |
| [6] | Johnson, S.L., Cuellar, A. and Gershon, A. (2016) The Influence of Trauma, Life Events, and Social Relationships on Bipolar Depression. Psychiatric Clinics of North America, 39, 87-94. https://doi.org/10.1016/j.psc.2015.09.003 |
| [7] | Akiskal, H.S. and McKinney, W.T. (1973) Depressive Disorders: Towards a Unified Hypothesis. Science, 182, 20-29. https://doi.org/10.1126/science.182.4107.20 |
| [8] | Maletic, V. and Raison, C. (2014) Integrated Neurobiology of Bipolar Disorder. Front. Psychiatry, 5, 98. https://doi.org/10.3389/fpsyt.2014.00098 |
| [9] | Hasler, G. (2010) Pathophysiology of Depression: Do We Have Any Solid Evidence of Interest to Clinicians? World Psychiatry, 9, 155-161. https://doi.org/10.1002/j.2051-5545.2010.tb00298.x |
| [10] | Verkhratsky, A., Rodrigues, J.J. and Steardo, L. (2014) Astrogliopathy: A Central Element of Neuropsychiatric Diseases. Neuroscientist, 20, 576-588. https://doi.org/10.1177/1073858413510208 |
| [11] | Rial, D., Lemos, C., Pinheiro, H., Duarte, J., Goncalves, F.Q., et al. (2015) Depression as a Glial-Based Synaptic Dysfunction. Frontiers in Cellular Neuroscience, 9, 521. |
| [12] | Perea, G. and Arague, A. (2003) Tripartite Synapses: Bidirectional Communication between Astrocytes and Neurons. Journal of Physiology, 548, 15. |
| [13] | Halassa, M.M. and Haydon, P.G. (2010) Integrated Brain Circuits: Astrocytic Networks Modulate Neuronal Activity and Behavior. Annual Review of Physiology, 72, 335-355. https://doi.org/10.1146/annurev-physiol-021909-135843 |
| [14] | Mitterauer, B. (2004) Imbalance of Glial-Neuronal Interaction in synapses: a possible Mechanism of the Pathophysiology of Bipolar Disorder. Neuroscientist, 10, 199-206. https://doi.org/10.1177/107385403262248 |
| [15] | Mitterauer, B.J. (2009) Narziss and Echo. Ein psychobiologisches Modell der Depression. Springer, Vienna. https://doi.org/10.1007/978-3-211-99140-4 |
| [16] | Mitterauer, B.J. (2010) Synaptic Imbalances in Endogenous Psychoses. BioSystems, 100, 113-121. https://doi.org/10.1016/j.biosystems.2010.02.006 |
| [17] | Mitterauer, B.J. (2015) Balancing and Imbalancing Effects of Astrocytic Receptors in Tripartite Synapses. Common Pathophysiological Model of Mental Disorders and Epilepsy. Medical Hypotheses, 84, 315-320. https://doi.org/10.1016/j.mehy.2015.01.025 |
| [18] | Mitterauer, B.J. (2007) Where and How Could Intentional Programs Be Generated in the Brain? A Hypothetical Model Based on Glial-Neuronal Interactions. BioSystems, 88, 101-112. https://doi.org/10.1016/j.biosystems.2006.04.003 |
| [19] | Arague, A., Parpura, V., Sanzgiri, R.P. and Haydon, P.G. (1999) Tripartite Synapses: Glia, the Unacknowledged Partner. Trends in Neurosciences, 22, 208-215. https://doi.org/10.1016/S0166-2236(98)01349-6 |
| [20] | Mitterauer, B.J. (2006) Pseudo Omnipotence: A Model of the Manic Syndrome. Nova Science Publishers, New York, 161-178. |
| [21] | Bibring, E. (1953) The Mechanism of Depression. In: Greenacre, P., Ed., Affective Disorders, International Universities Press, New York, 13-48. |
| [22] | Champagne, F.A. and Mashoodt, R. (2012) Genes in Context: Gene-Environment Interplay and Origins of Individual Differences in Behavior. Current Directions in Psychological Science, 18, 127-131. https://doi.org/10.1111/j.1467-8721.2009.01622.x |
| [23] | Bagot, R.C. and Meaney, M.J. (2010) Epigenetics and the Biological Basis of Gene x Environment Interactions. Journal of American Academy of Child and Adolescent Psychiatry, 49, 752-771. https://doi.org/10.1016/j.jaac.2010.06.001 |
| [24] | Stuffrein-Roberts, S., Joyce, P.R. and Kennedy, M.A. (2008) Role of Epigenetics in Mental Disorders. The Australian and New Zealand Journal of Psychiatry, 42, 97-107. https://doi.org/10.1080/00048670701787495 |
| [25] | Dalton, V.S., Kolshus, E. and McLaughlin, D.M. (2014) Epigenetics and Depression: Return of the Repressed. Journal of Affective Disorders, 155, 1-12. https://doi.org/10.1016/j.jad.2013.10.028 |
| [26] | Freud, S. (1917) Trauer und Melancholie. In: Fischer, S., Ed., Gesammelte Werke, Bd 11, Frankfurt. |
| [27] | Ovidius Naso (1993) Metamorphosen. Artemis, München. |
| [28] | Ransom, B.R. and Giaume, C. (2013) Gap Junctions, Hemichannels. In: Kettenman, H. and Rason, B.R., Eds., Neuroglia, Oxford University Press, New York, 292-305. |
| [29] | Guenther, G. (1963) Das Bewußtsein der Maschinen. Agis, Krefeld. |
| [30] | Mitterauer, B.J. (2014) Pathophysiology of Schizophrenia Based on Impaired Glial-Neuronal Interactions. Open Journal of Medical Psychology, 3, 126-140. https://doi.org/10.4236/ojmp.2014.32016 |
| [31] | Mitterauer, B.J. (2011) Downregulation and Upregulation of Glial Connexin May Cause Synaptic Imbalances Responsible for the Pathophysiology of Bipolar Disorder. CNS Neuroscience and Therapeutics, 17, 281-293. https://doi.org/10.1111/j.1755-5949.2010.00178.x |
| [32] | Quesseveur, G., Portal, B., Basic, J.A., et al. (2015) Attenuated Levels of Hippocampal Connexin 43 and Its Phosphorylation Correlate with Antidepressant- and Anxiolytic-Like Activation in Mice. Frontiers in Cellular Neuroscience, 9, 490. |
| [33] | Solan, J.L. and Lampe, P.D. (2009) Connexin 43 Phosphorylation: Structural Changes and Biological Effects. Biochemical Journal, 419, 261-272. https://doi.org/10.1042/BJ20082319 |
| [34] | Mayberg, H.S., Brannan, S.K., Tekell, J.L, et al. (2000) Regional Metabolic Effects of Fluoxetine in Major Depression: Serial Changes and Relationship to Clinical Response. Biological Psychiatry, 48, 830-843. https://doi.org/10.1016/S0006-3223(00)01036-2 |
| [35] | Kettenmann, H. and Zorec, R. (2013) Release of Gliotransmitters and Transmitter Receptors in Astrocytes. In: Kettenmann, H. and Ransom, B.R., Eds., Neuroglia, Oxford University Press, New York, 197-211. https://doi.org/10.1093/med/9780199794591.001.0001 |
| [36] | Yu, W., Mechavar, N., Kratic, S., et al. (2012) Upregulation of Astrocytic Alpha 7 Nicotinic Receptors in Alzheimer’s Disease Brain—Possibly Relevant to Amyloid Pathology. Molecular Neurodegeneration, 7, 7. http://www.molecularneurodegeneration.com/content/7/S1/07 |
| [37] | Joshida, Y., Nagai, A., Kobayashi, S. and Kein, S.U. (2006) Upregulation of Protease-Activated Receptor-1 in Astrocytes in Parkinson’s Disease: Astrocyte-Mediated Neuroprotection through Levels of Glutathione Peroxidase. Journal of Neuropathology & Experimental Neurology, 65, 66-77. https://doi.org/10.1097/01.jnen.0000195941.48033.eb |
| [38] | Borroto-Escuela, D.O. and Fuxe, K. (2017) Diversity and Bias through Dopamine D2R Heteroreceptor Complexes. Current Opinion in Pharmacology, 32, 16-22. https://doi.org/10.1016/j.coph.2016.10.004 |
| [39] | Muner, A. (2017) Mixed States in Bipolar Disorder: Etiology, Pathogenesis and Treatment. Chonnam Medical Journal, 53, 1-13. https://doi.org/10.4068/cmj.2017.53.1.1 |
| [40] | Milior, G., Lecours, C., Samson, L., et al. (2015) Fractalkine Receptor Deficiency Impairs Microglial and Neuronal Responsiveness to Chronic Stress. Brain, Behavior, and Immunity, 55, 114-125. |
| [41] | Berk, M., Dodd, S., Kauer-Sant’anna, et al. (2007) Dopamine Dysregulation Syndrome: Implications for a Dopamine Hypothesis of Bipolar Disorder. Acta Psychiatrica Scandinavica, No. 434, 41-49. https://doi.org/10.1111/j.1600-0447.2007.01058.x |
| [42] | Goncales, P. (2009) Behavior Modes, Pathways and Overall Trajectories: Eigenvector and Eigenvalue Analysis of Dynamic Systems. System Dynamics Review, 25, 35-62. https://doi.org/10.1002/sdr.414 |
| [43] | McCulloch, W.S. (1965) Embodiments of Mind. Cambridge University Press, Cambridge. |
| [44] | Humphries, M.D., Gurney, K. and Prescott, T.J. (2006) The Brainstem Reticular Formation Is a Small-World, Not Scale-Free, Network. Proceedings of the Royal Society B, 273, 503-511. https://doi.org/10.1098/rspb.2005.3354 |
| [45] | Okada, Y., Sasaki, T., Oku, Y., et al. (2012) Preinspiratory Calcium Rise in Putative Pre-Bötzinger Complex Astrocytes. The Journal of Physiology, 590, 4933-4944. |
| [46] | Marquette, P., Vendier, D., Kodala, A., et al. (2015) An Astrocyte-Dependent Mechanism of Neuronal Rhythmogenesis. Nature Neuroscience, 18, 844-854. https://doi.org/10.1038/nn.4013 |
| [47] | Barbas, H. and Zikopoulos, B. (2007) The Prefrontal Cortex and Flexible Behavior. Neuroscientist, 13, 532-545. https://doi.org/10.1177/1073858407301369 |
| [48] | Mitterauer, B.J. (2015) Model of the Reticular Formation of the Brainstem. Based on Glial-Neuronal Interactions. Cognitive Computation, 7, 64-73. https://doi.org/10.1007/s12559-014-9260-5 |
| [49] | Giaume, C. and Naus, C.C. (2013) Connexins, Gap Junctions, and Glia. WIREs Membrane Transport and Signaling, 2, 133-142. |
| [50] | Kettenmann, H., Kirchhoff, F. and Verkhratsky, A. (2012) Microglia: New Notes for the Synaptic Stripper. Neuron, 77, 10-18. |
| [51] | Munkholm, K., Vinberg, M. and Vedel, K.L. (2013) Cytokines in Bipolar Disorder: A Systematic Review and Meta-Analysis. Journal of Affective Disorders, 144, 16-27. https://doi.org/10.1016/j.jad.2012.06.010 |
| [52] | Jakobsson, J., Bjerke, M., Sahebi, S., et al. (2015) Monocyte and Microglial Activation in Patients with Mood-Stabilized Bipolar Disorder. Journal of Psychiatry & Neuroscience, 40, 250-258. https://doi.org/10.1503/jpn.140183 |
| [53] | Gigante, A.D., Young, L.T., Yatham, L.N., et al. (2011) Monophometric Post-Mortem Studies in Bipolar Disorder: Possible Association with Oxidative Stress and Apoptosis. International Journal of Neuropharmacology, 14, 1075-1086. |
| [54] | Carter, C.J. (2007) Multiple Genes and Factors Associated with Bipolar Disorder Converge on Growth Factor and Stress Activated Kinase Pathways Controlling Translation Initiation: Implications for Oligodendrocyte Viability. Neurochemistry International, 50, 461-490. |
| [55] | Chen, C.H., Suckling, J., Lennox, B.R., et al. (2011) A Quantitative Meta-Analysis of fMRI Studies in Bipolar Disorder. Bipolar Disorders, 13, 1-15. https://doi.org/10.1111/j.1399-5618.2011.00893.x |
| [56] | Martinowich, K., Schloesser, R.J. and Manji, H.K. (2009) Bipolar Disorder: From Genes to Behavior Pathways. The Journal of Clinical Investigation, 119, 726-736. https://doi.org/10.1172/JCI37703 |
| [57] | Pritz, W.F. and Mitterauer, B.J. (1984) Bipolar Mood Disorders: An Affected Sibling Study. Psychopathology, 17, 67-79. https://doi.org/10.1159/000284006 |
| [58] | Mitterauer, B. (2000) Clock Genes, Feedback Loops and Their Possible Role in the Etiology of Bipolar Disorders: An Integrative Model. Medical Hypotheses, 55, 155-159. https://doi.org/10.1054/mehy.1999.1039 |
| [59] | Bunney, W.E. and Bunney, B.G. (2000) Molecular Clock Genes in Man and Lower Animals: Possible Implications for Circadian Abnormalities in Depression. Neuropsychopharmacology, 22, 335-345. https://doi.org/10.1016/S0893-133X(99)00145-1 |
| [60] | Milhiet, V., Etain, B., Boudebesse, C. and Bellivier, F. (2011) Circadian Biomarkers, Circadian Genes and Bipolar Disorders. Journal of Physiology, 105, 183-910. https://doi.org/10.1016/j.jphysparis.2011.07.002 |
| [61] | Craddock, N. and Sklar, P. (2013) Genetics of Bipolar Disorder. The Lancet, 381, 1654-1662. https://doi.org/10.1016/S0140-6736(13)60855-7 |
| [62] | Gonzalez, R. (2014) The Relationship between Bipolar Disorder and Biological Rhythms. Journal of Clinical Psychiatry, 75, e323-e331. https://doi.org/10.4088/JCP.13r08507 |
| [63] | Losi, G., Mariotti, L., Sessolo, M. and Camignoto, G. (2017) New Tools to Study Astrocyte Ca2+ Signal Dynamics in Brain Networks in Vivo. Frontiers in Cellular Neuroscience, 11, 134. https://doi.org/10.3389/fncel.2017.00134 |
| [64] | Robertson, J.M. (2013) Astrocyte Domains and the Three-Dimensional and Seamless Expression of Consciousness and Explicit Memories. Medical Hypothesis, 81, 1017-1024. https://doi.org/10.1016/j.mehy.2013.09.021 |
| [65] | Geddes, J.R. and Miklowitz, D. (2013) Treatment of Bipolar Disorder. The Lancet, 381. https://doi.org/10.1016/S0140-6736(13)60857-0 |
